 科技服务
科技服务工作动态
RNA病毒编码的依赖RNA的RNA聚合酶(RNA-dependent RNA polymerase,简称RdRP)是一类独特的核酸聚合酶,在病毒基因组复制和转录过程中发挥核心作用,是抗病毒药物研究的热点靶标。病毒的RdRP由于可与其他功能域融合或与其他病毒蛋白共折叠,其整体结构多样性较高,但其催化核心区的三维结构则较为保守,因此RdRP兼具多样性和保守性。由于RNA病毒的宿主范围几乎囊括了所有细胞形式的生命体,在与不同宿主的共同进化过程中,RdRP作为RNA病毒最保守的蛋白,其与病毒的宿主适应性之间的关联并不清晰。
黄病毒包括乙型脑炎病毒(Japanese encephalitis virus,简称JEV)、登革病毒(dengue virus,简称DENV)、寨卡病毒、蜱传脑炎病毒(tick-borne encephalitis virus,简称TBEV)等多种人类致病病原,是一类分布广泛、种类繁多的单股正链RNA病毒,与丙型肝炎病毒和经典猪瘟病毒同属于黄病毒科。黄病毒大多由吸血节肢动物如蚊和蜱作为媒介传播,可引起人类脑炎或出血性疾病,对人类健康构成重大威胁。黄病毒的RdRP位于病毒编码的非结构蛋白NS5的羧基端,与氨基端的甲基转移酶(MTase)形成天然融合体。此前蚊传黄病毒的NS5已有多个三维结构报道,包括全长蛋白、MTase区和RdRP区的结构,而蜱传黄病毒NS5蛋白的全长结构及RdRP三维结构则未获解析。
中国科学院武汉病毒研究所龚鹏研究员团队长期从事病毒RdRP的催化与调控机制研究,该团队此前分别解析了JEV和DENV的NS5全长蛋白晶体结构(Lu and Gong, PLoS Pathog 2013; Wu et al., PLoS Pathog 2020),并与武汉病毒所张波研究员团队合作,系统性揭示了NS5的构象多样性和保守性以及MTase调控RdRP的分子机制(Li et al. PLoS Negl Trop Dis 2014; Wu, et al. J Virol 2015; Wu et al., PLoS Pathog 2020)。该团队近期解析了分辨率为1.9 埃的TBEV MTase晶体结构(PDB号7D6M,图1A)和分辨率为3.2埃的TBEV RdRP的晶体结构(PDB号7D6N,图1B),获得了首个蜱传黄病毒RdRP的三维结构信息。通过黄病毒RdRP的序列分析以及与结构已知的蚊传黄病毒RdRP的结构进行比较,发现在RdRP保守的催化基序(motif )B和C之间存在一个值得关注的区域(以下称B-C连接区)。该区域位于RdRP手掌区的底部且暴露于蛋白表面,且在黄病毒属的不同宿主分类中具有明显的宿主相关多样性(图1B及图2)。
该团队设计了TBEV和JEV病毒间B-C连接区的替换突变,在酶学水平证实突变对RdRP催化功能不构成本质影响(图3,A-B)。团队通过与武汉病毒所王汉中研究员/郑振华研究员团队和张波研究员团队合作,在细胞水平分别评测了突变对TBEV和JEV的影响,结果表明在两种病毒体系中突变后的病毒不能维持在细胞中的增殖(图3,C-D)。这些结果提示B-C连接区很可能参与了RdRP催化以外与病毒增殖相关的重要过程。通过对正链、负链和双链RNA病毒中的代表性RdRP的B-C连接区进行结构与序列的系统分析,在RNA病毒大家族中进一步发现该区域在结构和序列长度方面具有较高的多样性(图4),提示RdRP的B-C连接区很可能是RNA病毒共有的一个宿主适应热点区域,此项研究为病毒RdRP的调控机制研究及RdRP相关的宿主适应研究提供了重要线索。
此项研究主要受到国家重点研发计划项目“畜禽重要病原共感染与协同致病机制研究”(2018YFD0500100,项目负责人为中国农业科学院上海兽医研究所丁铲研究员)和NSFC面上项目(31670154; 32070185)的支持。博士研究生杨婕妤和博士后景旭平为论文的共同第一作者,主要完成了结构与酶学研究工作,龚鹏、郑振华和张波为共同通讯作者,王汉中/郑振华团队的实验师易文富、硕士研究生姚琛和张波团队的博士后李晓丹分别完成了TBEV和JEV病毒学研究工作,相关论文近期于Nucleic Acids Research(《核酸研究》)上在线发表,原文链接为: https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkaa1250/6066638?guestAccessKey=c6d13ee0-287e-4d7b-97dc-01d9456423e7
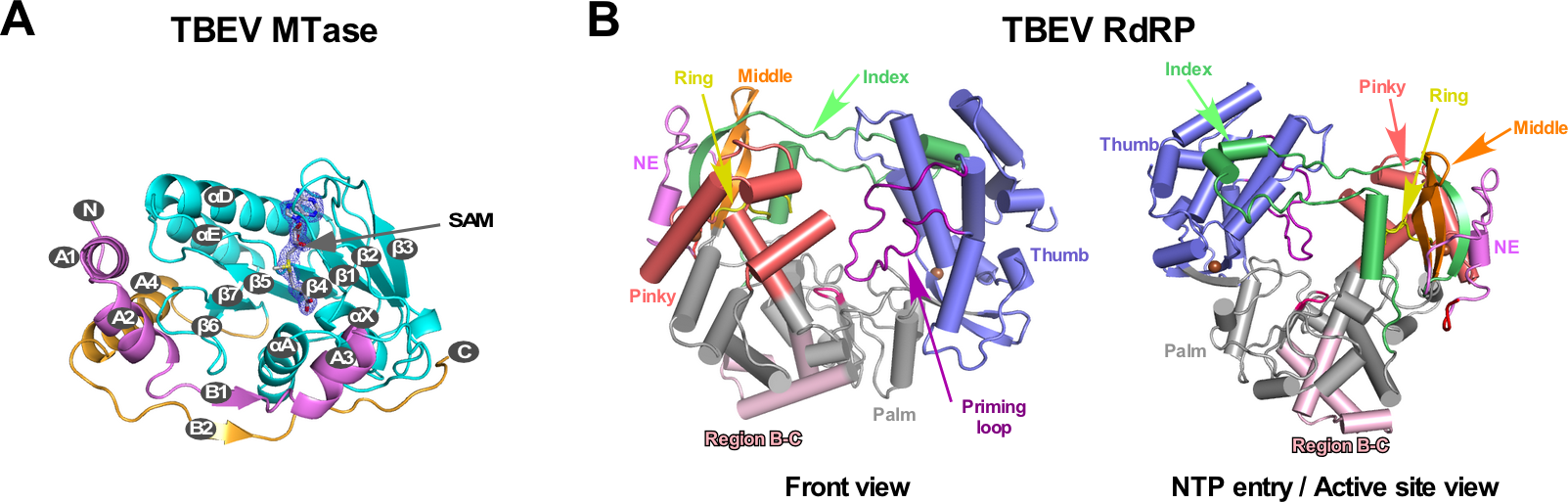
图1. TBEV MTase和RdRP的晶体结构。A)MTase结构中结合了一个甲基供体SAM分子。B)RdRP由拇指、手指和手掌三个结构域组成,B-C连接区(浅粉色)位于手掌区底部。
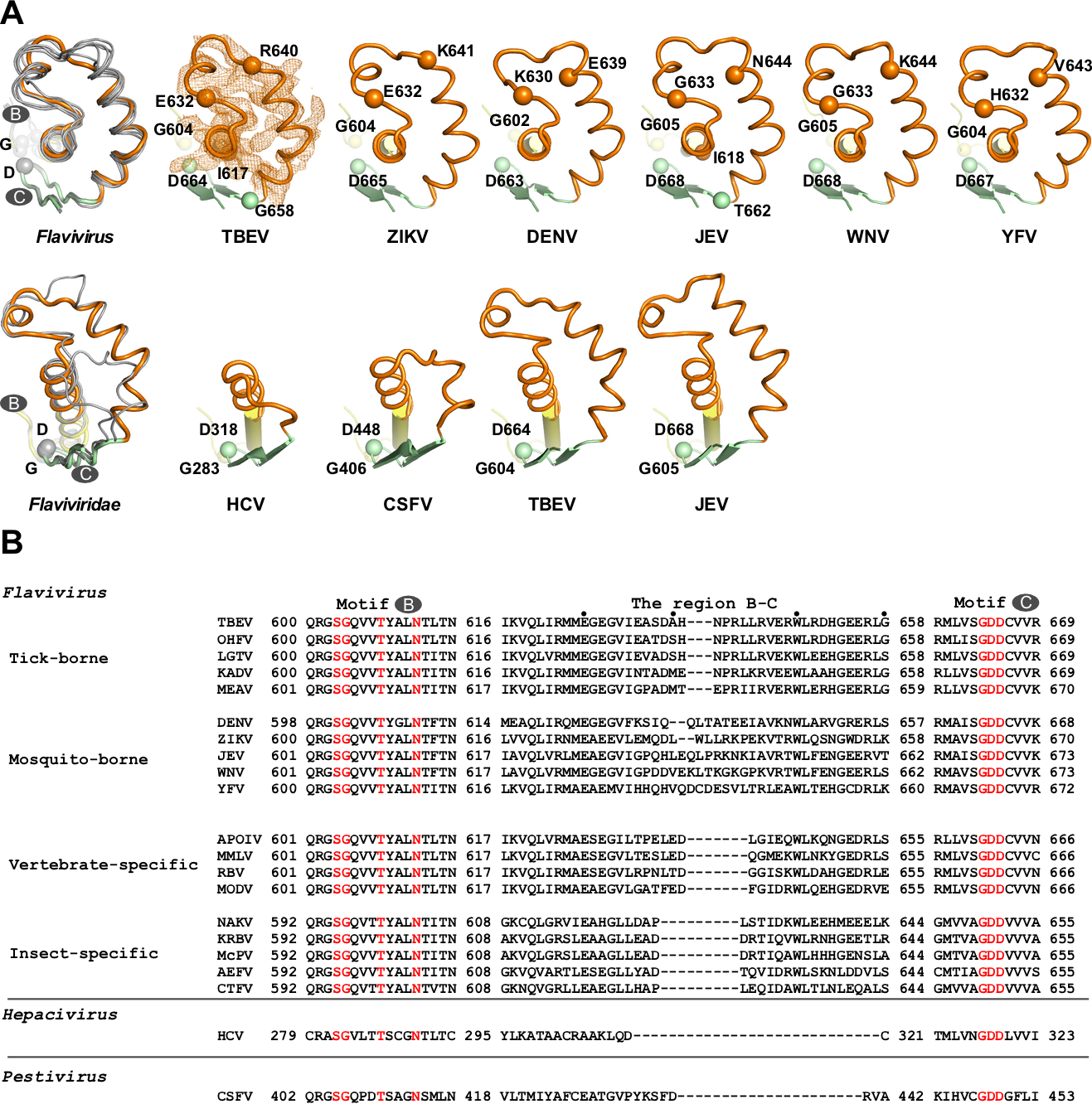
图2. 黄病毒科RdRPs中B-C连接区结构和序列的多样性。A)第一排中TBEV属于蜱传黄病毒,其余属于蚊传黄病毒。第二排中TBEV和JEV属于黄病毒属,而HCV和CSFV分别属于黄病毒科中的肝炎病毒属和瘟病毒属。其中motif B、B-C连接区和motif C分别用黄色、橙色和浅绿色表示。B)黄病毒NS5中B-C区域的序列比对。根据宿主的不同,黄病毒可以分为蚊传、蜱传、仅在昆虫中传播和仅在脊椎动物中传播四类。
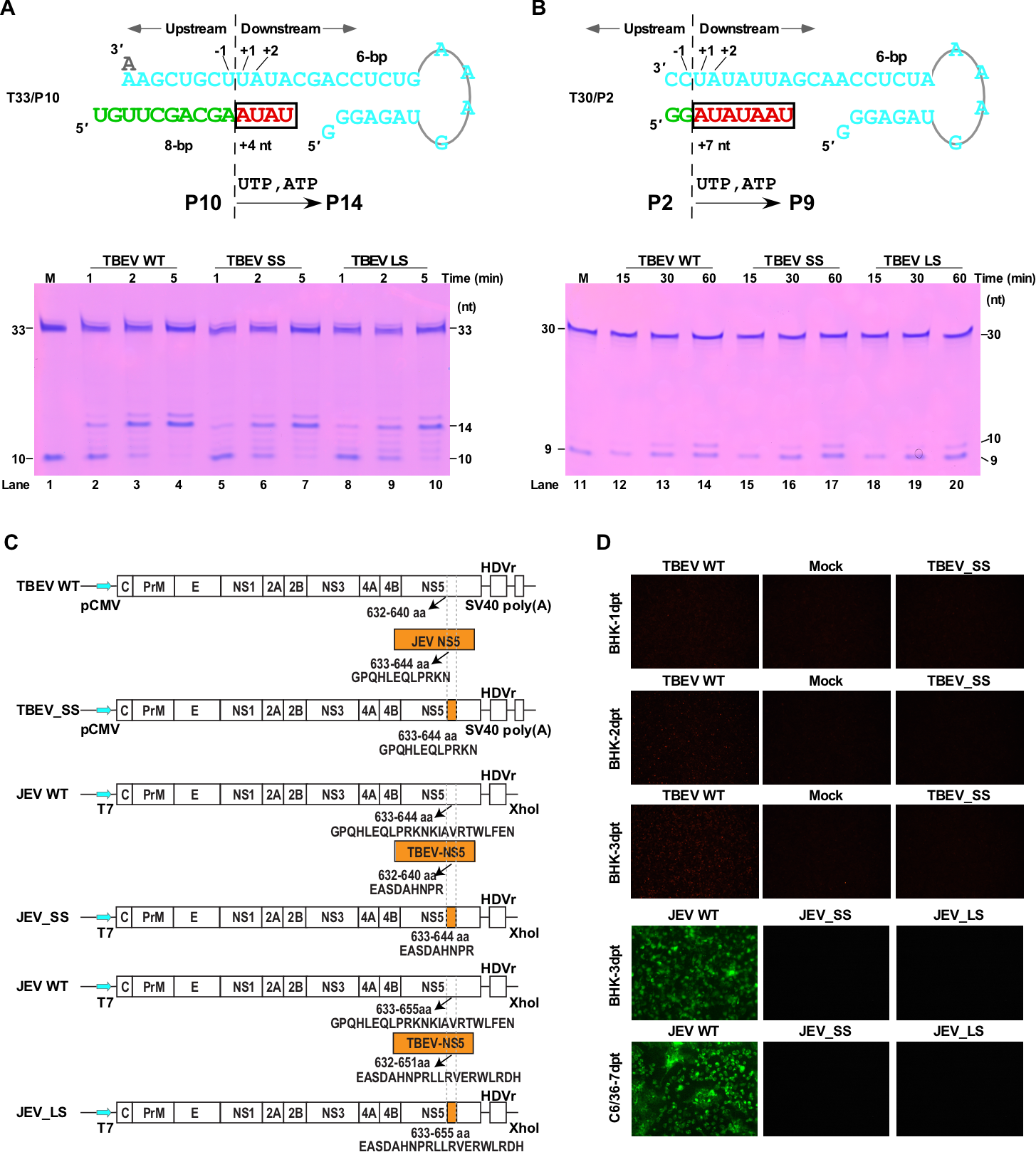
图3. 病毒种间B-C连接区替换突变影响病毒增殖。A-B)将连接区替换为JEV序列的TBEV NS5表现出与野生型聚合酶相似的依赖引物(A)和从头合成(B)活性。C-D)在细胞水平上TBEV和JEV的连接区替换突变体均不能实现病毒增殖。
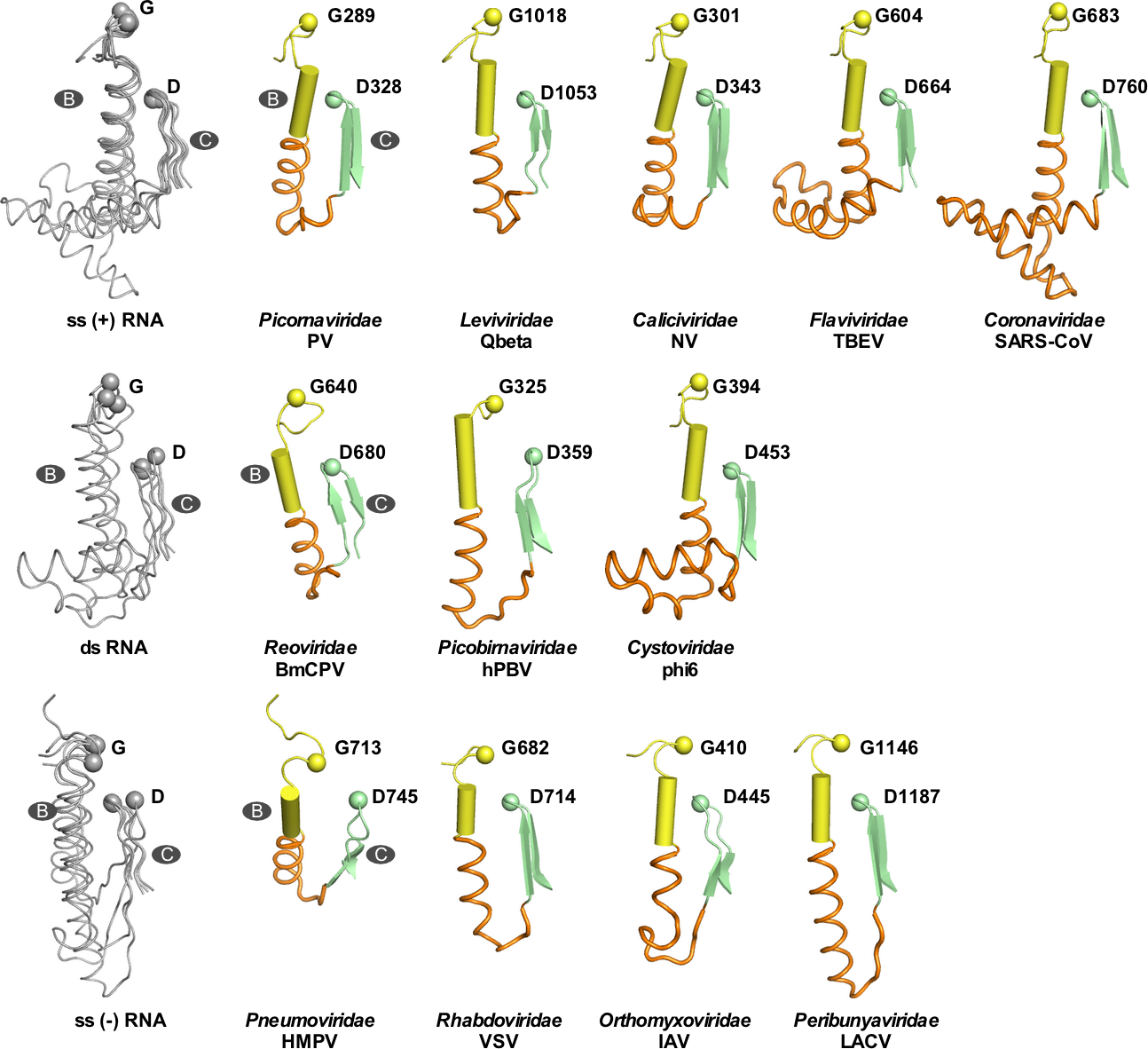
图4. 病毒RdRP的B-C连接区展现出较高的多样性。上中下三行分别为正链、负链和双链RNA病毒代表性RdRP的连接区结构。每个彩色结构分别代表一个RNA病毒科。其中B-C连接区用橙色显示。左侧为多结构叠印图(灰色)。